Существует несколько типов классификации процесса дробления.
По характеру образования и расположению бластомеров:
Полное (голобластическое) - характерно для зигот, содержащих мало желтка (мезо- и изолецитальные яйца), при этом борозды дробления проходят через все яйцо, а имеющийся у них желток включается в вегетативные бластомеры;
Неполное (меробластическое) - характерно для зигот, содержащих большие запасы белков желтка (полилецитальные яйца), при этом борозды дробления не проникают в богатую желтком область цитоплазмы.
В зависимости от размеров образовавшихся бластомеров:
равномерное - бластомеры на анимальном и вегетативном полюсе имеют одинаковые размеры;
неравномерное - на анимальном полюсе сосредоточены более мелкие бластомеры, чем на вегетативном.
По скорости формирования бластомеров:
синхронное - при одинаковой скорости образования бластомеров на обоих полюсах зиготы;
асинхронное - на анимальном полюсе скорость образования бластомеров выше, чем на вегетативном.
Выделяют четыре основных типа голобластического дробления . Данная классификация основана на взаимном пространственном расположении бластомеров:
Радиальное;
Спиральное;
Билатерально-симметричное;
Неправильное (анархическое).
Радиальный тип дробления присущ голобластическим хордовым (ланцетник, круглоротые, осетровые рыбы, амфибии), иглокожим и некоторым другим группам.
При этом типе дробления бластомеры разных широтных ярусов располагаются, по крайней мере на ранних стадиях, довольно точно один над другим, так что полярная ось яйца служит осью поворотной симметрии.
Радиальный равномерный тип дробления характерен для яиц иглокожих (рис. 23).
У яйца лягушки наблюдается радиальный неравномерный тип дробления. Борозда первого деления дробления еще не завершила разделения богатой желтком цитоплазмы вегетативного полушария, а борозды второго деления уже закладываются вблизи от анимального полюса. Из-за большой концентрации желтка в вегетативной области борозды третьего деления дробления располагаются значительно ближе к анимальному полюсу (рис. 24).
В результате возникают область быстро делящихся бластомеров вблизи анимального полюса и область более медленно делящихся бластомеров вегетативного полюса.
Спиральный тип дробления характеризуется утерей элементов симметрии уже на стадии четырех, а иногда и двух бластомеров и присущ беспозвоночным (моллюски, кольчатые и ресничные черви), объединяемым в группу Spiralia.
Свое название этот тип дробления получил из-за того, что при взгляде с анимального полюса последовательно отделяющиеся четверки (квартеты) бластомеров поворачиваются относительно анимально-вегетативной оси то в правую, то в левую сторону, как бы образуя при наложении друг на друга спираль (рис. 25).
Знак спирального дробления, его дексио-(право-) или лео-(лево-) тропность, т. е. «закрученность», определяется геномом матери данной особи. Оно во многом отличается от радиального типа дробления.
Во-первых, яйца не делятся параллельно или перпендикулярно анимально-вегетативной оси. Плоскости делений дробления ориентированы наклонно, что приводит к спиральному расположению дочерних бластомеров.
Во-вторых, число контактов между клетками больше, чем при радиальном дроблении. В-третьих, зародыши со спиральным типом дробления проходят меньше делений до начала гаструляции. Возникающие таким образом бластулы обычно не имеют бластоцели (стерробластула).
Билатеральный тип дробления (круглые черви, оболочники) характеризуется наличием одной плоскости симметрии. Наиболее примечательная особенность этого типа дробления заключается в том, что плоскость первого деления устанавливает единственную плоскость симметрии зародыша (рис. 26).
Каждое последующее деление ориентируется по отношению к этой плоскости симметрии так, что половина зародыша по одну сторону от первой борозды представляет собой зеркальное отражение половины зародыша по другую ее сторону.
рис. 27. Анархическое дробление (по Токину, 1987)
При билатеральном типе дробления формируется одна плоскость симметрии: первая борозда проходит экваториально, далее анимальный бластомер делится меридиональной бороздой, а вегетативный - широтной. В результате получается Т - образная фигура из четырех бластомеров, не обладающая поворотной симметрией.
Путем поворота вегетативной пары бластомеров Т-образная фигура преобразуется в ромбическую. Этот поворот происходит в промежутке между делениями, в интерфазе.
При этом они могут распадаться, например под ударами волн, но из отдельных участков образуются полноценные зародыши. В результате плотного объединения бластомеров друг с другом в конце дробления образуется морула.
Основными типами меробластического дробления являются:
Поверхностное;
Дискоидальное.
При поверхностном дроблении после слияния пронуклеусов ядро зиготы делится на много ядер, которые с небольшим количеством цитоплазмы по цитоплазматическим мостикам переходят во внешний слой свободной от желтка цитоплазмы (периплазму) и равномерно там распределяются
(речь идет о центролецитальных яйцеклетках). Здесь ядра еще несколько раз синхронно делятся, располагаясь довольно близко друг к другу (рис. 28).
На этой стадии, еще до возникновения клеточных перегородок (так называемой синцитиальной бластодермы), ядра окружаются особыми структурами из микротрубочек, затем деление ядер становится асинхронным, между ними формируются клеточные перегородки и образуется базальная мембрана, отделяющая периплазму от центральной массы желтка. Борозды дробления появляются, но они не заходят глубоко в яйцо. Возникший поверхностный слой клеток называется клеточной бластодермой . Этот тип дробления характерен для большинства насекомых.
Первые две борозды проходят перпендикулярно друг другу, но далее строгий порядок прохождения борозд нарушается. При этом на бластомеры делится лишь тонкий диск цитоплазмы (бластодиск), расположенный на анимальном полюсе.
Зигота
Зигота (греч. zygote соединенная в пару) - диплоидная (содержащая полный двойной набор хромосом) клетка, образующаяся в результате оплодотворения (слияния яйцеклетки и сперматозоида). Зигота является тотипотентной (то есть, способной породить любую другую) клеткой. Термин ввёл немецкий ботаник Э. Страсбургер.
У человека первое митотическое деление зиготы происходит спустя примерно 30 часов после оплодотворения, что обусловлено сложными процессами подготовки к первому акту дробления. Клетки, образовавшиеся в результате дробления зиготы называют бластомерами. Первые деления зиготы называют "дроблениями" потому, что клетка именно дробится: дочерние клетки после каждого деления становятся всё мельче, а между делениями отсутствует стадия клеточного роста.
Развитие зиготы Зигота либо непосредственно после оплодотворения приступает к развитию, либо одевается плотной оболочкой и на некоторое время превращается в покоящуюся спору (часто называется зигоспорой) - характерно для многих грибов и водорослей.
Дробление
Период эмбрионального развития многоклеточного животного начинается с дробления зиготы и завершается рождением новой особи. Процесс дробления заключается в серии последовательных митотических делений зиготы. Образующиеся в результате нового деления зиготы две клетки и все последующие поколения клеток на этом этапе носят название бластомеров. В ходе дробления одно деление следует за другим, и не происходит роста образующихся бластомеров, вследствие чего каждое новое поколение бластомеров представлено более мелкими клетками. Эта особенность клеточных делений при развитии оплодотворенной яйцеклетки и определила появление образного термина - дробление зиготы.
У разных видов животных яйцеклетки различаются по количеству и характеру распределения в цитоплазме запасных питательных веществ (желтка). Это в значительной степени определяет характер последующего дробления зиготы. При небольшом количестве и равномерном распределении желтка в цитоплазме происходит деление всей массы зиготы с образованием одинаковых бластомеров - полное равномерное дробление (например, у млекопитающих). При скоплении желтка преимущественно у одного из полюсов зиготы происходит неравномерное дробление - образуются бластомеры, различающиеся по размерам: более крупные макромеры и микромеры (например, у амфибий). Если же яйцеклетка очень богата желтком, то дробится ее часть, свободная от желтка. Так, у пресмыкающихся, птиц дроблению подвергается лишь дисковидный участок зиготы у одного из полюсов, где располагается ядро - неполное, дискоидальное дробление. Наконец, у насекомых в процессе дробления задействован лишь поверхностный слой цитоплазмы зиготы - неполное, поверхностное дробление.
В результате дробления (когда число делящихся бластомеров достигает значительного числа) образуется бластула. В типичном случае (например, у ланцетника) бластула представляет собой полый шар, стенка которого образована одним слоем клеток (бластодерма). Полость бластулы - бластоцелъ, иначе называемая первичной полостью тела, заполнена жидкостью. У амфибий бластула имеет очень небольшую полость, а у некоторых животных (например, членистоногих) бластоцель может полностью отсутствовать.
Образуется зигота, способная к дальнейшему развитию. Деление зиготы называют дроблением. Дробление – это многократное деление зиготы после оплодотворения, в результате которого образуется многоклеточный зародыш.
Зигота делится очень быстро, клетки уменьшаются в размерах и не успевают расти. Поэтому зародыш не увеличивается в объеме. Клетки, образующиеся в результате , называются бластомерами, а перетяжки, отделяющие их друг от друга, называются бороздами дробления.
По направлению различают следующие борозды дробления: меридиональные – это борозды, которые делят зиготу от анимального к вегетативному полюсу; экваториальная борозда разделяет зиготу по экватору; широтные борозды проходят параллельно экваториальной борозде; тангенциальные борозды проходят параллельно поверхности зиготы.
Экваториальная борозда всегда одна, а меридиональных, широтных и тангенциальных может быть много. Направление борозд дробления всегда определяется положением веретена деления.
Дробление всегда проходит по определенным правилам:
Первое правило отражает местоположение веретена дробления в бластомере, а именно:
– веретено дробления располагается в сторону наибольшей протяженности цитоплазмы, свободной от включений.
Второе правило отражает направление борозд дробления:
– борозды дробления проходят всегда перпендикулярно веретену деления.
Третье правило отражает скорость прохождения борозд дробления:
– скорость прохождения борозд дробления обратно пропорционально количеству желтка в яйцеклетке, т.е. в той части клетки, где желтка мало, борозды будут проходить с большей скоростью, а в той части, где желтка больше, скорость прохождения борозд дробления замедляется.
Дробление зависит от количества и местоположения желтка в яйцеклетке. При небольшом содержании желтка дробится вся зигота, при значительном количестве дробится только часть зиготы, свободная от желтка. В связи с этим яйцеклетки разделяют на голобластические (дробящиеся полностью) и меробластические (с частичным дроблением). Следовательно, дробление зависит от количества желтка и с учетом ряда признаков подразделяется: по полноте охвата процессом материала зиготы на полное и неполное; по отношению размеров образующихся бластомеров на равномерное и неравномерное и по согласованности делений бластомеров – синхронное и асинхронное.
Полное дробление может быть равномерным и неравномерным. Полное равномерное характерно для яйцеклеток с небольшим количеством желтка и его более или менее равномерным расположением в . Таким типом дробится яйцеклетка . В этом случае первая борозда проходит от анимального к вегетативному полюсу, образуется два бластомера; вторая борозда тоже меридиональная, но проходит перпендикулярно первой, образуются четыре бластомера. Третья – экваториальная, образуются восемь бластомеров. После этого идет чередование меридиональных и широтных борозд дробления. Количество бластомеров после каждого деления увеличивается кратно двум (2; 4; 16; 32 и т.д.). В результате такого дробления образуется шарообразный зародыш, который называется бластулой
. Клетки, которые образуют стенку бластулы, называют бластодермой, а полость внутри бластоцелью. Анимальная часть бластулы называется – крышей, а вегетативная часть – дном бластулы.

Полное неравномерное дробление характерно для яйцеклеток со средним содержанием желтка, расположенным в вегетативной части. Такие яйцеклетки характерны для круглоротых и . При этом типе дробления образуются бластомеры неодинаковых размеров. В анимальном полюсе образуются мелкие бластомеры, которые называются микромерами, а в вегетативном – крупные – макромеры. Первые две борозды, как и у ланцетника, проходят меридионально; третья борозда соответствует экваториальной борозде, но сдвинута от экватора к анимальному полюсу. Поскольку в анимальном полюсе находится свободная от желтка цитоплазма, то здесь дробление происходит быстрее и образуются мелкие бластомеры. В вегетативном полюсе содержится основная масса желтка, поэтому борозды дробления проходят медленнее и образуются крупные бластомеры.
Неполное дробление характерно для телолецитальных и центролецитальных яйцеклеток. В дроблении принимает участие только часть яйца, свободная от желтка. Неполное дробление делится на дискоидальное (костистые , пресмыкающиеся, птицы) и поверхностное (членистоногие).
Неполным дискоидальным дроблением делятся телолецитальные яйцеклетки, у которых большое количество желтка сконцентрировано в вегетативной части. У этих яйцеклеток безжелтковая часть цитоплазмы в виде зародышевого диска распластана на желтке в анимальном полюсе. Дробление происходит только в области зародышевого диска. Вегетативная часть яйцеклетки, заполненная желтком, участия в дроблении не принимает. Толщина зародышевого диска незначительна, поэтому веретена дробления при первых четырех делениях располагаются горизонтально, а борозды дробления проходят вертикально. Образуется один ряд клеток. После нескольких делений клетки становятся высокими и веретена дробления располагаются в них в вертикальном направлении, а борозды дробления проходят параллельно поверхности яйца. В результате зародышевый диск превращается в пластинку, состоящую из нескольких рядов клеток. Между зародышевым диском и желтком возникает небольшая полость в виде щели, которая аналогична бластоцели.
Неполное поверхностное дробление наблюдается в центролецитальных яйцеклетках с большим количеством желтка в его середине. Цитоплазма в таких яйцеклетках располагается по периферии и незначительная ее часть в центре около ядра. Вся остальная часть клетки заполнена желтком. Через массу желтка проходят тонкие цитоплазматические тяжи, соединяющие периферическую цитоплазму с околоядерной. Дробление начинается с деления ядер, в результате количество ядер увеличивается. Они окружаются тонким ободком цитоплазмы, передвигаются к периферии и располагаются в свободной от желтка цитоплазме. Как только ядра попадают в поверхностный слой, он делится соответственно их количеству на бластомеры. В результате такого дробления вся центральная часть цитоплазмы перемещается к поверхности и сливается с периферической. Снаружи образуется сплошная бластодерма, из которой развивается зародыш, а внутри находится желток. Поверхностное дробление свойственно яйцеклеткам членистоногих.
На характер дробления оказывают влияние и свойства цитоплазмы, которые определяют взаимное расположение бластомеров. По этому признаку выделяют радиальное, спиральное и билатеральное дробление. При радиальном дроблении каждый верхний бластомер располагается точно под нижним (кишечнополостные, иглокожие, ланцетник и др.). При спиральном дроблении каждый верхний бластомер смещен относительно нижнего наполовину, т.е. каждый верхний бластомер располагается между двумя нижними. В этом случае бластомеры располагаются как бы по спирали (черви, моллюски). При билатеральном дроблении через зиготу можно провести только одну плоскость, по обеим сторонам которой будут наблюдаться одинаковые бластомеры (круглые черви, асцидии).
Антропология и концепции биологии Курчанов Николай Анатольевич
Стадии зиготы и дробления
Стадии зиготы и дробления
Зигота , образующаяся в результате слияния гамет, представляет собой одноклеточную стадию развития многоклеточного организма. Хотя продолжительность этой стадии обычно невелика, удается проследить происходящие в ней цитоморфологические и биохимические изменения. Эти изменения играют важную роль для последующих процессов эмбриогенеза. У ряда животных уже в зиготе начинается синтез белка на и-РНК, образованной еще во время оогенеза.
Дробление представляет собой процесс следующих друг за другом митотических делений, часто неравномерных. Клетки, образующиеся в процессе дробления, называются бластомерами . Заканчивается дробление образованием бластулы , обычно имеющей внутреннюю полость – бластоцель . Характерная особенность периода дробления – отсутствие роста. Хотя зародыш на стадии бластулы может состоять из сотен клеток, его размер не превышает размера зиготы.
В зависимости от типа яиц существует несколько видов дробления и бластул. Выделяют два основных типа яиц.
Гомолецитальные яйца – имеют расположенное в центре ядро и равномерно распределенный в цитоплазме желток.
Телолецитальные яйца – имеют четко выраженную полярность, эксцентричное расположение ядра и неравномерно распределенный в цитоплазме желток.
Гомолецитальные яйца дают обычно бластулы с одинаковыми бластомерами: целобластулу (с полостью) или морулу (без полости). Телолецитальные яйца дают бластулы с неравными бластомерами: амфибластулу (полное дробление) или дискобластулу (частичное дробление). У млекопитающих в результате дробления образуется морула, но затем в ходе полного асинхронного деления бластомеров возникает дополнительная стадия – зародышевой пузырек, или бластоциста .
Из книги Биология [Полный справочник для подготовки к ЕГЭ] автора Лернер Георгий Исаакович Из книги Основы психофизиологии автора Александров Юрий Из книги Теория адекватного питания и трофология [таблицы текстом] автора1.2. Стадии фиксации памяти Гипотеза о двух последовательно развивающихся следах. Согласно гипотезе, формирование энграммы осуществляется в два этапа: первый характеризуется неустойчивой формой следа и существует в течение непродолжительного периода. Это этап
Из книги Теория адекватного питания и трофология [таблицы картинками] автора Уголев Александр Михайлович3. СТАДИИ МЕДЛЕННОГО СНА И БЫСТРЫЙ СОН Основные данные, полученные за годы многочисленных и разнообразных исследований сна, сводятся к следующему. Сон – не перерыв в деятельности мозга, это просто иное состояние. Во время сна мозг проходит через несколько различных фаз,
Из книги Антропология и концепции биологии автора Курчанов Николай Анатольевич Из книги автора9.2. Происхождение и ранние стадии развития жизни Современная теория происхождения жизни - это теория появления структур, которые, развиваясь и усложняясь, приобретали определенные функциональные свойства. По-видимому, эти свойства могли возникнуть уже на ранних этапах
Из книги автораСтадии фотосинтеза В процессе фотосинтеза различают две стадии – световую и темновую.Во время световой фазы фотосинтеза энергия Солнца используется для синтеза АТФ и высокоэнергетических переносчиков электронов. Световая энергия, поглощенная любой молекулой
После сближения женского и мужского пронуклеусов, которое продолжается у млекопитающих около 12 ч, образуется зигота - одноклеточный зародыш. Уже на стадии зиготы выявляются презумптивные зоны (лат. presumptio - вероятность, предположение) как источники развития соответствующих участков бластулы, из которых в дальнейшем формируются зародышевые листки.
Рис. Зигота человека в стадии сближения мужского и женского ядер (пронуклеусов): (по Б.П.Хватову).
1 - женское ядро; 2 - мужское ядро.
Дробление и образование бластулы
Дробление - последовательное митотическое деление зиготы на клетки (бластомеры) без роста дочерних клеток до размеров материнской.
Образующиеся бластомеры остаются объединенными в единый организм зародыша. В зиготе образуется митотическое веретено между отдаляющимися к полюсам центриолями, внесенными сперматозоидом. Пронуклеусы вступают в стадию профазы с формированием объединенного диплоидного набора (Метод выявления презумптивных зон предложен немецким эмбриологом Фогтом) хромосом яйцеклетки и сперматозоида. Пройдя все остальные фазы митотического деления, зигота разделяется на две дочерние клетки - бластомеры. Вследствие фактического отсутствия G 1 -периода, во время которого происходит рост клеток, образовавшихся в результате деления, клетки гораздо меньше материнской, поэтому и величина зародыша в целом в этот период независимо от числа составляющих его клеток не превышает величину исходной клетки - зиготы. Все это позволило назвать описываемый процесс дроблением (т.е. измельчением), а клетки, образующиеся в процессе дробления, - бластомерами.
Дробление зиготы человека начинается к концу первых суток и характеризуется как полное неравномерное асинхронное. В течение первых суток оно происходит медленно. Первое дробление (деление) зиготы завершается через 30 ч, в результате образуется 2 бластомера, по- крытых оболочкой оплодотворения. За стадией двух бластомеров следует стадия трех бластомеров.
С первых же дроблений зиготы формируются два вида бластомеров - «темные» и «светлые». «Светлые», более мелкие, бластомеры дробятся быстрее и располагаются одним слоем вокруг крупных «темных», которые оказываются в середине зародыша. Из поверхностных «светлых» бластомеров в дальнейшем возникает трофобласт, связывающий зародыш с материнским организмом и обеспечивающий его питание. Внутренние, «темные», бластомеры формируют эмбриобласт, из которого образуются тело зародыша и некоторые внезародышевые органы (амнион, желточный мешок, аллантоис).
Начиная с трех суток, дробление идет быстрее, и на 4-е сутки зародыш состоит из 7-12 бластомеров. Уже через 50-60 ч образуется плотное скопление клеток - морула, а на 3-4-е сутки начинается формирование бластоцисты - полого пузырька, заполненного жидкостью (рис).

Рис. Зародыш человека на ранних стадиях развития (по Гертигу и Рокку).
А - стадия двух бластомеров; Б - бластоциста; I - эмбриобласт, 2 - трофобласт; 3 - полость бласто цисты.
Бластоциста в течение 3 сут перемещается по яйцеводу к матке и через 4 сут. попадает в матку. Бластоциста находится в полости матки в свободном виде в течение 2 дней (5-е и б-е сутки), и эта стадия обозначается как свободная бластоциста. К этому времени бластоциста увеличивается благодаря росту числа бластомеров - клеток эмбриобласта и трофобласта - до 100 и более вследствие усиленного всасывания трофобластом секрета маточных желез, а также вследствие активной выработки жидкости самим трофобластом (рис).
Эмбриобласт располагается в виде узелка зародышевых клеток («зародышевый узелок»), который прикрепляется изнутри к трофобласту на одном из полюсов бластоцисты и начинается имплантация.
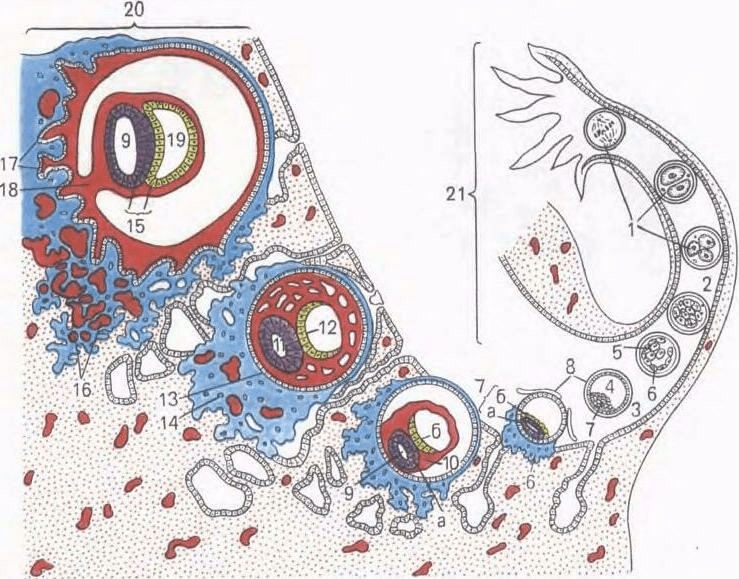
Рис. 37. Дробление, гаструляция и имплантация зародыша человека (схема).
1 - дробление, 2 - морула; 3 - бластоциста; 4 - полость бластоцисты; 5 - эмбриобласт; 6 - трофобласт; 7 - зародышевый узелок: а - эпибласт, б - гипобласт: 8 - оболочка оплодотворения; 9 - амниотический (эктодермальный) пузырек; 10 - внезародышевая мезодерма; II - эктодерма; 12 - энтодерма; 13 - цитотрофобласт; 14 - симпластотрофобласт; 15 - зародышевый диск; 16 - лакуны с материнской кровью; 17 - хорион; 18 - амниотическая ножка; 19 - желточный пузырек; 20 - слизистая оболочка матки; 21 - яйцевод.






