Hay varios tipos de clasificación del proceso de trituración.
Por la naturaleza de la formación y la ubicación de los blastómeros:
Completo (holoblástico) - característico de los cigotos que contienen poca yema (huevos meso- e isolecíticos), mientras que los surcos de división atraviesan todo el huevo, y la yema que tienen está incluida en los blastómeros vegetativos;
Incompleto (meroblástico): característico de los cigotos que contienen grandes reservas de proteínas de la yema (huevos polilecitales), mientras que los surcos de trituración no penetran en el citoplasma rico en yema.
Según el tamaño de los blastómeros formados:
uniforme- los blastómeros de los polos animal y vegetativo son del mismo tamaño;
desigual- Los blastómeros más pequeños se concentran en el polo animal que en el vegetativo.
Según la tasa de formación de blastómeros:
sincrónico- al mismo ritmo de formación de blastómeros en ambos polos del cigoto;
asincrónico- en el polo animal, la tasa de formación de blastómeros es mayor que en el vegetativo.
Asignar cuatro tipos principales de fragmentación holoblástica. Esta clasificación se basa en la disposición espacial mutua de los blastómeros:
Radial;
Espiral;
Simetría bilateral;
Mal (anarquista).
El tipo radial de aplastamiento es inherente a los cordados holoblásticos (lancetas, ciclóstomos, esturiones, anfibios), equinodermos y algunos otros grupos.
En este tipo de división, los blastómeros de diferentes etapas latitudinales se ubican, al menos en las primeras etapas, exactamente uno encima del otro, de modo que el eje polar del huevo sirve como eje de simetría rotacional.
El tipo de aplastamiento uniforme radial es característico de los huevos de equinodermo (Fig. 23).
En un huevo de rana, se observa un tipo de aplastamiento radial desigual. El surco de la primera división aún no ha completado la división del citoplasma rico en yema del hemisferio vegetativo, y los surcos de la segunda división ya se están formando cerca del polo animal. Debido a la alta concentración de yema en la región vegetativa, los surcos de la tercera división se ubican mucho más cerca del polo animal (Fig. 24).
Como resultado, surge un área de blastómeros de división rápida cerca del polo animal y un área de blastómeros de división más lenta del polo vegetativo.
El tipo de trituración en espiral se caracteriza por la pérdida de elementos de simetría ya en la etapa de cuatro, ya veces dos blastómeros, y es inherente a los invertebrados (moluscos, anélidos y gusanos ciliares), que se unen en el grupo Spiralia.
Este tipo de fragmentación recibió su nombre debido al hecho de que, cuando se ve desde el polo animal, los cuádruples (cuartetos) de blastómeros que se separan sucesivamente giran en relación con el eje animal-vegetativo hacia la derecha o hacia la izquierda, como si formaran una espiral cuando superpuestos entre sí (Fig. .25).
El signo de la fragmentación en espiral, su tropismo dexio-(derecha-) o leo-(izquierda-), es decir, "giro", está determinado por el genoma de la madre de un individuo determinado. Difiere en muchos aspectos del tipo de trituración radial.
Primero, los huevos no se dividen en forma paralela o perpendicular al eje animal-vegetativo. Los planos de las divisiones de clivaje están orientados oblicuamente, lo que da lugar a una disposición en espiral de los blastómeros hijos.
En segundo lugar, el número de contactos entre celdas es mayor que con la trituración radial. En tercer lugar, los embriones con un tipo de hendidura en espiral experimentan menos divisiones antes del inicio de la gastrulación. Las blástulas resultantes generalmente no tienen blastocele (esterroblastula).
El tipo de trituración bilateral (gusanos redondos, tunicados) se caracteriza por la presencia de un plano de simetría. La característica más notable de este tipo de división es que el plano de la primera división establece el único plano de simetría del núcleo (Fig. 26).
Cada división subsiguiente se orienta con respecto a este plano de simetría de manera que la mitad del embrión de un lado del primer surco es una imagen especular de la mitad del embrión del otro lado.
arroz. 27. Fragmentación anarquista (según Tokin, 1987)
Con el tipo de trituración bilateral, se forma un plano de simetría: el primer surco corre ecuatorialmente, luego el blastómero animal se divide por el surco meridional y el blastómero vegetativo se divide por el latitudinal. El resultado es una figura en forma de T de cuatro blastómeros, que no tiene simetría rotacional.
Al girar el par vegetativo de blastómeros, la figura en forma de T se transforma en una rómbica. Esta rotación ocurre en el intervalo entre divisiones, en la interfase.
Al mismo tiempo, pueden desintegrarse, por ejemplo, bajo el impacto de las olas, pero se forman embriones de pleno derecho a partir de secciones individuales. Como resultado de la densa asociación de blastómeros entre sí, al final de la trituración, un mórula.
Los principales tipos de fragmentación meroblástica son:
superficial;
Discoidal.
Durante el aplastamiento superficial después de la fusión de los pronúcleos, el núcleo del cigoto se divide en muchos núcleos que, con una pequeña cantidad de citoplasma, pasan a través de los puentes citoplasmáticos hacia la capa externa del citoplasma libre de yema (periplasma) y se distribuyen uniformemente. allí.
(Estamos hablando de ovocitos centrolecíticos). Aquí, los núcleos se dividen varias veces más sincrónicamente, estando ubicados bastante cerca uno del otro (Fig. 28).
En esta etapa, incluso antes de la aparición de los tabiques celulares (el llamado blastodermo sincitial), los núcleos están rodeados por estructuras especiales de microtúbulos, luego la división nuclear se vuelve asincrónica, se forman tabiques celulares entre ellos y se forma una membrana basal que separa el periplasma de la masa central de la yema. Aparecen surcos de división, pero no penetran profundamente en el huevo. La capa superficial resultante de células se denomina capa celular. blastodermo. Este tipo de aplastamiento es característico de la mayoría de los insectos.
Los dos primeros surcos corren perpendiculares entre sí, pero luego se viola el orden estricto de los surcos. En este caso, solo un disco delgado del citoplasma (blastodisco), ubicado en el polo animal, se divide en blastómeros.
Cigoto
El cigoto (del griego cigoto emparejado) es una célula diploide (que contiene un conjunto doble completo de cromosomas) que resulta de la fertilización (la fusión de un óvulo y un espermatozoide). El cigoto es una célula totipotente (es decir, capaz de producir cualquier otra). El término fue introducido por el botánico alemán E. Strasburger.
En humanos, la primera división mitótica del cigoto se produce aproximadamente 30 horas después de la fecundación, lo que se debe a los complejos procesos de preparación para el primer acto de trituración. Las células formadas como resultado de la trituración del cigoto se denominan blastómeros. Las primeras divisiones del cigoto se denominan "aplastamiento" porque la célula se aplasta: después de cada división, las células hijas se vuelven cada vez más pequeñas y no hay una etapa de crecimiento celular entre divisiones.
Desarrollo del cigoto El cigoto comienza a desarrollarse inmediatamente después de la fecundación o se reviste de una capa densa y durante algún tiempo se convierte en una espora en reposo (a menudo llamada cigospora), típica de muchos hongos y algas.
División
El período de desarrollo embrionario de un animal multicelular comienza con la fragmentación del cigoto y termina con el nacimiento de un nuevo individuo. El proceso de escisión consiste en una serie de sucesivas divisiones mitóticas del cigoto. Las dos células formadas como resultado de una nueva división del cigoto y todas las generaciones posteriores de células en esta etapa se denominan blastómeros. Durante la trituración, una división sigue a otra y los blastómeros resultantes no crecen, por lo que cada nueva generación de blastómeros está representada por células más pequeñas. Esta característica de las divisiones celulares durante el desarrollo de un óvulo fertilizado determinó la aparición de un término figurativo: aplastamiento del cigoto.
En diferentes especies animales, los huevos difieren en la cantidad y la naturaleza de la distribución de los nutrientes de reserva (yema) en el citoplasma. Esto determina en gran medida la naturaleza de la posterior fragmentación del cigoto. Con una pequeña cantidad y distribución uniforme de la yema en el citoplasma, toda la masa del cigoto se divide con la formación de blastómeros idénticos: trituración uniforme completa (por ejemplo, en mamíferos). Cuando la yema se acumula predominantemente en uno de los polos del cigoto, se produce una fragmentación desigual: se forman blastómeros que difieren en tamaño: macrómeros y micrómeros más grandes (por ejemplo, en anfibios). Si el huevo es muy rico en yema, entonces se tritura su parte libre de yema. Entonces, en los reptiles, las aves, solo la sección en forma de disco del cigoto en uno de los polos, donde se encuentra el núcleo, sufre fragmentación: fragmentación discoidal incompleta. Finalmente, en los insectos, solo la capa superficial del citoplasma del cigoto está involucrada en el proceso de trituración: trituración incompleta y superficial.
Como resultado del aplastamiento (cuando el número de blastómeros en división alcanza un número significativo), se forma una blástula. En un caso típico (por ejemplo, en la lanceta), la blástula es una bola hueca, cuya pared está formada por una sola capa de células (blastodermo). La cavidad de la blástula - blastocoel, también llamada cavidad corporal primaria, está llena de líquido. En los anfibios, la blástula tiene una cavidad muy pequeña y en algunos animales (como los artrópodos), el blastocele puede estar completamente ausente.
Se forma un cigoto, capaz de un mayor desarrollo. La división de un cigoto se llama clivaje. División- Esta es la división repetida del cigoto después de la fertilización, como resultado de lo cual se forma un embrión multicelular.
El cigoto se divide muy rápidamente, las células disminuyen de tamaño y no tienen tiempo de crecer. Por lo tanto, el embrión no aumenta de volumen. Las células resultantes se denominan blastómeros y las constricciones que las separan se denominan surcos de división.
Los siguientes surcos de trituración se distinguen en la dirección: meridional: son surcos que dividen el cigoto del animal al polo vegetativo; el surco ecuatorial divide el cigoto a lo largo del ecuador; los surcos latitudinales corren paralelos al surco ecuatorial; los surcos tangenciales corren paralelos a la superficie del cigoto.
El surco ecuatorial es siempre uno, pero puede haber muchos surcos meridionales, latitudinales y tangenciales. La dirección de los surcos de trituración siempre está determinada por la posición del husillo de división.
La trituración siempre se lleva a cabo de acuerdo con ciertas reglas:
La primera regla refleja la ubicación del huso de clivaje en el blastómero, a saber:
- el huso de clivaje se ubica en la dirección de mayor extensión del citoplasma, libre de inclusiones.
La segunda regla refleja la dirección de los surcos de trituración:
- los surcos de trituración discurren siempre perpendiculares al huso de fisión.
La tercera regla refleja la velocidad de trituración de surcos:
- la velocidad de paso de los surcos de división es inversamente proporcional a la cantidad de yema en el huevo, es decir en aquella parte de la celda donde hay poca yema, los surcos pasarán a mayor velocidad, y en la parte donde hay más yema, la velocidad de paso de los surcos de clivaje se hace más lenta.
La escisión depende de la cantidad y la ubicación de la yema en el huevo. Con una pequeña cantidad de yema, se tritura todo el cigoto, con una cantidad significativa, solo se tritura una parte del cigoto libre de yema. En este sentido, los huevos se dividen en holoblásticos (completamente triturados) y meroblásticos (con trituración parcial). En consecuencia, la trituración depende de la cantidad de yema y, teniendo en cuenta una serie de características, se subdivide: según la totalidad de la cobertura del material del cigoto por el proceso, en completo e incompleto; de acuerdo con la proporción de los tamaños de los blastómeros formados a uniforme y desigual, y de acuerdo con la consistencia de las divisiones de blastómeros: sincrónicas y asincrónicas.
La trituración completa puede ser uniforme y desigual. La uniformidad completa es característica de los huevos con una pequeña cantidad de yema y su disposición más o menos uniforme en. Este tipo de huevo triturado. En este caso, el primer surco va desde el animal hasta el polo vegetativo, se forman dos blastómeros; el segundo surco también es meridional, pero corre perpendicular al primero, se forman cuatro blastómeros. El tercero es ecuatorial, se forman ocho blastómeros. Después de esto, hay una alternancia de surcos de trituración meridionales y latitudinales. El número de blastómeros después de cada división aumenta por un factor de dos (2; 4; 16; 32, etc.). Como resultado de tal trituración, se forma un embrión esférico, que se llama blástula. Las células que forman la pared de la blástula se llaman blastodermo, y la cavidad interior se llama blastocele. La parte animal de la blástula se llama techo y la parte vegetal se llama parte inferior de la blástula.

La trituración desigual completa es típica de los huevos con un contenido promedio de yema ubicado en la parte vegetativa. Tales huevos son característicos de cyclostomes y. Donde tipo de aplastamiento Se forman blastómeros de tamaños desiguales. En el polo animal se forman pequeños blastómeros, que se denominan micrómeros, y en el polo vegetativo, grandes, macrómeros. Los dos primeros surcos, como los de la lanceta, discurren meridionalmente; el tercer surco corresponde al surco ecuatorial, pero está desplazado del ecuador al polo animal. Dado que el citoplasma libre de yema se encuentra en el polo animal, aquí la fragmentación ocurre más rápido y se forman pequeños blastómeros. El polo vegetativo contiene la mayor parte de la yema, por lo que los surcos de división pasan más lentamente y se forman grandes blastómeros.
La escisión incompleta es característica de los ovocitos telolecíticos y centrolecíticos. Solo la parte del huevo que no tiene yema participa en la trituración. El aplastamiento incompleto se divide en discoidal (óseo, reptiles, aves) y superficial (artrópodos).
Los ovocitos telolecíticos se dividen por escisión discoidal incompleta, en la que una gran cantidad de yema se concentra en la parte vegetativa. En estos huevos, la parte libre de yema del citoplasma en forma de disco germinal se extiende sobre la yema en el polo animal. La escisión ocurre solo en la región del disco germinal. La parte vegetativa del huevo, llena de yema, no participa en la trituración. El grosor del disco germinal es insignificante, por lo que los husos de división en las primeras cuatro divisiones son horizontales y los surcos de división corren verticalmente. Se forma una fila de celdas. Después de varias divisiones, las células se vuelven altas y los husos de división se ubican en dirección vertical, y los surcos de división corren paralelos a la superficie del huevo. Como resultado, el disco germinal se convierte en una placa que consta de varias filas de células. Entre el disco germinal y la yema aparece una pequeña cavidad en forma de hendidura, similar al blastocele.
Se observa aplastamiento superficial incompleto en huevos centrolecíticos con gran cantidad de yema en su centro. El citoplasma de tales óvulos se encuentra a lo largo de la periferia y una pequeña parte se encuentra en el centro, cerca del núcleo. El resto de la celda se llena de yema. Finas hebras citoplasmáticas atraviesan la masa de la yema, conectando el citoplasma periférico con el perinuclear. La fragmentación comienza con la fisión de los núcleos, como resultado, aumenta el número de núcleos. Están rodeados por un borde delgado del citoplasma, se mueven hacia la periferia y se ubican en el citoplasma libre de yema. Tan pronto como los núcleos entran en la capa superficial, se divide en blastómeros según su número. Como resultado de tal aplastamiento, toda la parte central del citoplasma se mueve hacia la superficie y se fusiona con la periférica. En el exterior se forma un blastodermo continuo, a partir del cual se desarrolla el embrión, y en el interior está la yema. El aplastamiento superficial es característico de los huevos de artrópodos.
La naturaleza del aplastamiento también está influenciada por las propiedades del citoplasma, que determinan la posición relativa de los blastómeros. Sobre esta base, se distinguen el aplastamiento radial, espiral y bilateral. Con el aplastamiento radial, cada blastómero superior se ubica exactamente debajo del inferior (celenterados, equinodermos, lancetas, etc.). Durante la trituración en espiral, cada blastómero superior se desplaza a la mitad con respecto al inferior, es decir, cada blastómero superior se encuentra entre los dos inferiores. En este caso, los blastómeros están dispuestos como en espiral (gusanos, moluscos). Con el aplastamiento bilateral, solo se puede dibujar un plano a través del cigoto, en ambos lados se observarán blastómeros idénticos (gusanos redondos, ascidias).
Antropología y conceptos de biología Kurchanov Nikolai Anatolievich
Etapas de cigoto y escisión
Etapas de cigoto y escisión
Cigoto, formado como resultado de la fusión de gametos, es una etapa unicelular en el desarrollo de un organismo multicelular. Aunque la duración de esta etapa suele ser corta, es posible rastrear los cambios citomorfológicos y bioquímicos que ocurren en ella. Estos cambios juegan un papel importante en los procesos posteriores de embriogénesis. En varios animales, ya en el cigoto, la síntesis de proteínas comienza en el ARNm formado durante la ovogénesis.
División Es un proceso de sucesivas divisiones mitóticas, a menudo desiguales. Las células que se forman durante la escisión se llaman blastómeros. La trituración termina con la formación blástula, por lo general tiene una cavidad interna - blastocoel. Un rasgo característico del período de trituración es la falta de crecimiento. Aunque el embrión en la etapa de blástula puede constar de cientos de células, su tamaño no excede el tamaño del cigoto.
Según el tipo de óvulos, existen varios tipos de trituración y blástula. Hay dos tipos principales de huevos.
Huevos homolecíticos - tienen un núcleo situado en el centro y una yema distribuida uniformemente en el citoplasma.
huevos telolecíticos - tienen una polaridad pronunciada, una disposición excéntrica del núcleo y una yema desigualmente distribuida en el citoplasma.
Los huevos homolecíticos suelen dar lugar a blástulas con blastómeros idénticos: celoblastula(con cavidad) o morulú(sin cavidad). Los huevos telolecíticos dan lugar a blástulas con blastómeros desiguales: anfiblastula(aplastamiento completo) o discoblastula(aplastamiento parcial). En los mamíferos, como resultado del aplastamiento, se forma una mórula, pero luego, durante la división asincrónica completa de los blastómeros, ocurre una etapa adicional: vesícula germinal, o blastocisto.
Del libro Biología [Una guía completa para prepararse para el examen] autor Lerner Georgy Isaakovich Del libro Fundamentos de Psicofisiología autor Alexandrov Yuri Del libro The Theory of Adequate Nutrition and Trophology [tablas en el texto] autor1.2. Etapas de la fijación de la memoria Hipótesis de dos huellas que se desarrollan secuencialmente. Según la hipótesis, la formación del engrama se lleva a cabo en dos etapas: la primera se caracteriza por una forma inestable de la huella y existe por un período corto. este es el escenario
Del libro The Theory of Adequate Nutrition and Trophology [tablas con imágenes] autor Ugolev Alexander Mijailovich3. ETAPAS DEL SUEÑO NREM Y DEL SUEÑO REM Los principales datos obtenidos a lo largo de los años de numerosos y variados estudios sobre el sueño son los siguientes. El sueño no es una interrupción en la actividad del cerebro, es simplemente un estado diferente. Durante el sueño, el cerebro pasa por varias fases diferentes,
Del libro Antropología y Conceptos de Biología autor Kurchanov Nikolái Anatolievich Del libro del autor9.2. Origen y primeras etapas del desarrollo de la vida La teoría moderna del origen de la vida es la teoría de la aparición de estructuras que, desarrollándose y haciéndose más complejas, adquirieron ciertas propiedades funcionales. Aparentemente, estas propiedades podrían haber surgido ya en las primeras etapas.
Del libro del autorEtapas de la fotosíntesis En el proceso de la fotosíntesis, se distinguen dos etapas: clara y oscura.Durante la fase luminosa de la fotosíntesis, la energía del Sol se utiliza para sintetizar ATP y transportadores de electrones de alta energía. Energía luminosa absorbida por cualquier molécula.
Después de la convergencia de los pronúcleos femenino y masculino, que dura alrededor de 12 horas en los mamíferos, un cigoto- embrión unicelular. Ya en la etapa de cigoto, las zonas presuntivas (lat. presumptio - probabilidad, suposición) se revelan como fuentes de desarrollo de las secciones correspondientes de la blástula, a partir de las cuales se forman posteriormente las capas germinales.
Arroz. Cigoto humano en la etapa de convergencia de los núcleos masculino y femenino (pronúcleos): (según B.P. Khvatov).
1 - núcleo femenino; 2 - núcleo masculino.
Escisión y formación de la blástula.
Escisión: división mitótica secuencial del cigoto en células (blastómeros) sin el crecimiento de células hijas del tamaño de la madre.
Los blastómeros resultantes permanecen unidos en un solo organismo del embrión. En el cigoto se forma un huso mitótico entre los centriolos que se alejan hacia los polos, introducido por el espermatozoide. Los pronúcleos entran en la etapa de profase con la formación de un conjunto diploide combinado (el método para detectar zonas presuntivas fue propuesto por el embriólogo alemán Vogt) de cromosomas de óvulos y espermatozoides. Después de pasar por todas las demás fases de la división mitótica, el cigoto se divide en dos células hijas: los blastómeros. Debido a la ausencia virtual del período G 1, durante el cual crecen las células formadas como resultado de la división, las células son mucho más pequeñas que la célula madre, por lo tanto, el tamaño del embrión en su conjunto durante este período, independientemente de la número de sus células constituyentes, no excede el tamaño de la célula original - el cigoto. Todo esto hizo posible llamar al proceso descrito trituración (es decir, trituración) y las células formadas en el proceso de trituración: blastómeros.
La escisión del cigoto humano comienza al final del primer día y se caracteriza por ser asincrónica completamente desigual. Durante los primeros días, sucede lentamente. La primera escisión (división) del cigoto se completa después de 30 horas, dando como resultado la formación de 2 blastómeros cubiertos con una membrana de fertilización. La etapa de dos blastómeros es seguida por la etapa de tres blastómeros.
Desde la primera trituración del cigoto, se forman dos tipos de blastómeros: "oscuro" y "claro". Los blastómeros "claros", más pequeños, se trituran más rápido y se disponen en una capa alrededor de los grandes "oscuros", que se encuentran en el centro del embrión. De los blastómeros "ligeros" de la superficie, surge posteriormente un trofoblasto, que conecta el embrión con el cuerpo de la madre y proporciona su nutrición. Los blastómeros internos, "oscuros", forman el embrioblasto, a partir del cual se forman el cuerpo del embrión y algunos órganos extraembrionarios (amnios, saco vitelino, alantoides).
A partir de los tres días, la fragmentación avanza más rápido, y al cuarto día el embrión consta de 7-12 blastómeros. Después de 50-60 horas, se forma una densa acumulación de células, la mórula, y en el tercer o cuarto día comienza la formación de un blastocisto, una vesícula hueca llena de líquido (Fig.

Arroz. El embrión humano en las primeras etapas de desarrollo (según Hertig y Rock).
A - etapa de dos blastómeros; B - blastocisto; I - embrioblasto, 2 - trofoblasto; 3 - cavidad del blastoquiste.
El blastocisto se mueve a través del oviducto hacia el útero dentro de los 3 días y después de los 4 días. entra en el útero. El blastocisto está libre en la cavidad uterina durante 2 días (días 5 y 6), y esta etapa se denomina blastocisto libre. En este momento, el blastocisto aumenta debido a un aumento en el número de blastómeros (células de embrioblasto y trofoblasto) hasta 100 o más debido a una mayor absorción de la secreción de las glándulas uterinas por el trofoblasto, y también debido a la producción activa de fluido por el propio trofoblasto (Fig.
El embrioblasto se encuentra en forma de un paquete de células germinales ("paquete de gérmenes"), que se une desde el interior al trofoblasto en uno de los polos del blastocisto y comienza la implantación.
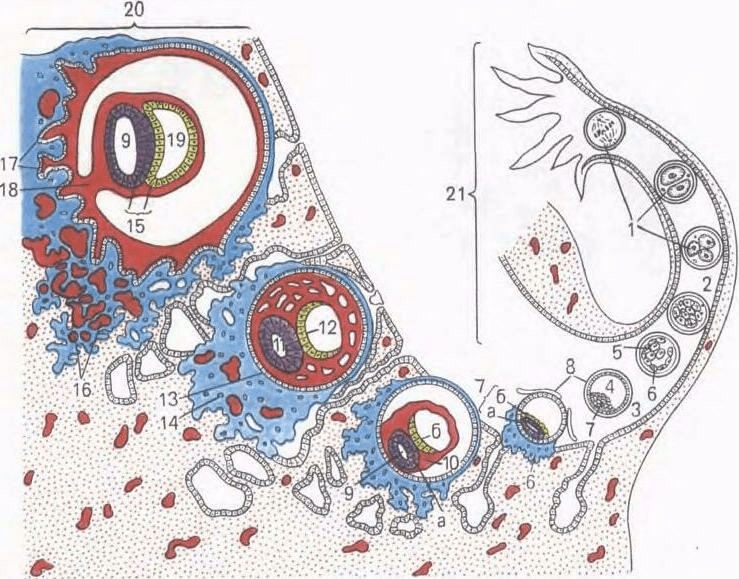
Arroz. 37. Escisión, gastrulación e implantación del embrión humano (esquema).
1 - trituración, 2 - mórula; 3 - blastocisto; 4 - cavidad de blastocisto; 5 - embrioblasto; 6 - trofoblasto; 7 - nódulo germinal: a - epiblasto, b - hipoblasto: 8 - membrana de fertilización; 9 - vesícula amniótica (ectodérmica); 10 - mesodermo extraembrionario; II - ectodermo; 12 - endodermo; 13 - citotrofoblasto; 14 - simplastotrofoblasto; 15 - disco germinal; 16 - lagunas con sangre materna; 17 - corion; 18 - pierna amniótica; 19 - vesícula de yema; 20 - membrana mucosa del útero; 21 - oviducto.






